ABSTRACT
The distribution of genetic variation in a species depends on both gene flow and population history. In this study, we demonstrate the utility of nested clade analysis for identifying historical factorsóignored by most traditional analysesóand provide one of the first studies to apply this approach to plants. PCR-RFLP markers were used to survey 912 bp of cpDNA in Delphinium nuttallii (Ranunculaceae), a native of the Pacific Northwest. Twelve populations located throughout the range of the species were examined, sampling five individuals per population. Sixteen polymorphisms and nine haplotypes were observed. A single most parsimonious network was found, which contained only one homoplastic trait. AMOVA, a non-historical analysis, suggested a regime of restricted gene flow to explain the pattern of genetic variation that was observed (F ST = 0.87, P < 0.0001). A simple historical approach, the exact nested contingency analysis, rejected the null hypothesis of no geographical association for all but one nesting group (P < 0.05 for these clades), but could not identify the source of these associations. The full nested clade analysis detected restricted gene flow or past fragmentation for all one-step clades, and contiguous range expansion at high cladogram levels. These inferences are supported by the biology of D. nuttallii which has no special adaptations for seed dispersal, and by the geologic history of the area. Post-Pleistocene glacial retreat would have allowed northward expansion of D. nuttallii during the last 15,000 years, and the Missoula Floods of 13,500-15,000 years ago may have permitted subsequent dispersal through the Columbia River Gorge into the western portion of its current range. For D. nuttallii, nested clade analysis detected and identified important historical factors that could not be resolved by simpler analyses employed in this study.Key words: chloroplast DNA, Delphinium nuttallii, nested clade analysis, phylogeography
INTRODUCTION
The distribution of genetic variation in a species depends on the effects of both gene flow and population history. Phylogeography has made great strides towards exploring these processes and cataloging the interactive effects of life history and biogeography, yet this approach still faces two main challenges: (1) statistical analyses that measure the role of gene flow are typically based on equilibrium models that exclude the possibility of recent historical events; (2) when interpretations implicating a role for population history (e.g. range expansion, fragmentation, long-distance colonization) are offered, they are usually supported by qualitative explanations of observed data rather than quantitative statistical tests. Nested clade analysis appears to offer a solution to both of these problems. 1,2 The approach incorporates temporal information evident in intraspecific haplotype phylogenies and uses a permutational approach to compare observed patterns of genetic variation to those predicted for gene flow and various historical scenarios. The present study uses a nested clade analysis to separate the effects of gene flow and population history for Delphinium nuttallii (Ranunculaceae), a plant endemic to the Northwestern region of the U.S. Delphinium nuttallii is well suited to this study because of known historical events in its range (Pleistocene glaciation and the Missoula Floods), and because high levels of chloroplast DNA (cpDNA) variation have previously been detected in the species by analyzing restriction fragment polymorphisms in PCR-amplified cpDNA regions (PCR-RFLPs). The results of our study demonstrate the utility of nested clade analysis for plant phylogeography, and provide insights into historical processes shaping intraspecific genetic variation in plants.
METHODS

Figure 1. Closeup of Delphinium nuttallii inflorescence. (photo by K. Karoly) |
|
Study Species. Delphinium nuttalliinutt (Ranunculaceae) is an herbaceous perennial that occupies a broad geographic and ecological range in the Northwest from mounded prairies in the Puget Sound lowlands, to oak woodlands (Quercus garryanna) found in association with basaltic soils in the Columbia River Gorge, to the high-elevation prairies of the eastern Cascade Mountain range (Figure 1). Plants flower in May - July and have an outcrossing mating system with protandrous flowers. Controlled crossing studies show that cpDNA inheritance in Delphinium is uniparental (maternal).3
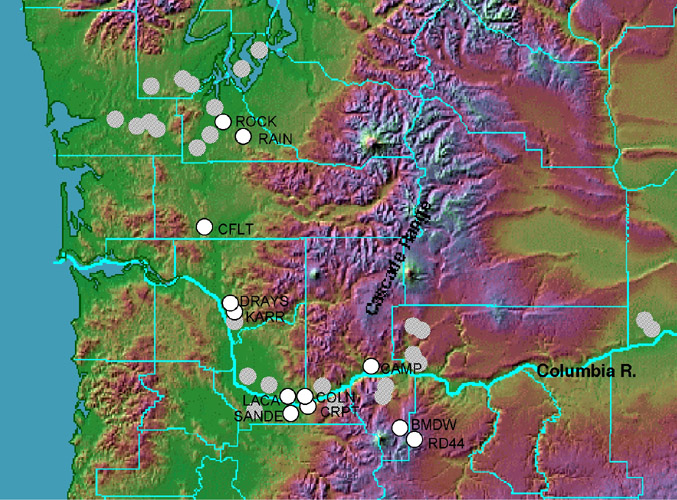
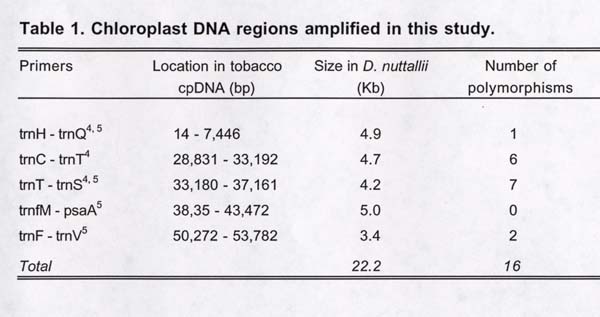
Data collection. Twelve populations located throughout the range of the species were examined, sampling five individuals each (N = 60; Figure 2). First, one plant sample from each population was screened for polymorphisms, using PCR-RFLPs. 4,5 Four amplified regions (18 kb total) were screened with nine 4-cutter restriction enzymes. All individuals were then genotyped using the eight polymorphic enzyme/primer combinations discovered in screening, plus eight additional enzyme/primer combinations previously shown to be polymorphic in D. nuttallii (Table 1).
Analysis. Resulting presence/absence and length variation data were used to construct a haplotype network based on maximum parsimony. A nesting diagram was then constructed from the network.6,7 The nested clade analysis and exact, nested contingency test were implemented using GeoDis 2.0 software.8 No outgroup weights were included and statistical significance was evaluated using 10,000 random permutations. Outputs were interpreted using an inference key.1 An analysis of molecular variance (AMOVA) was used to calculate F ST, and implemented with Arlequin 2.000, treating each population as a group. 9
|
RESULTS
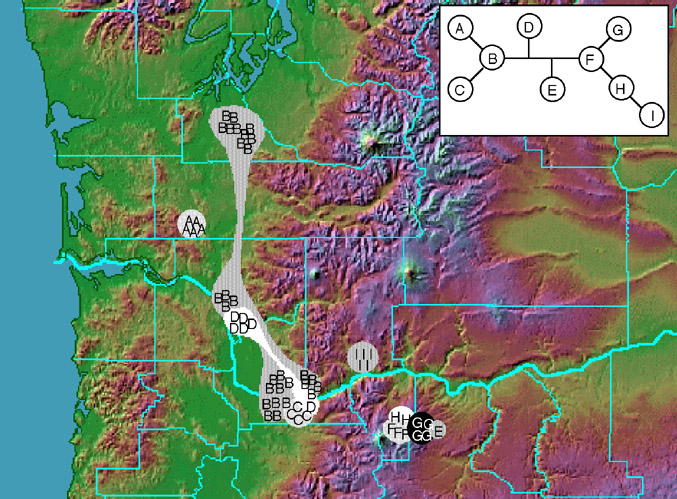
Figure 3. Haplotype distribution (haplotype network is inset). (Map modified from Color Landform Atlas of the United States, used with permission from R. Sterner) |
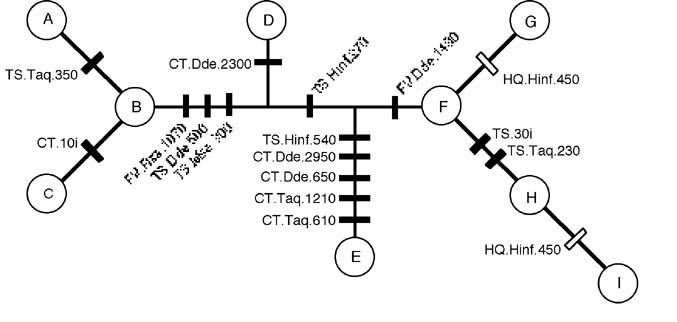
Figure 4. Chloroplast DNA haplotype network, with mutations (locus.enzyme.size) indicated along branches separating observed haplotypes. |
Extensive cpDNA variation was detected in Delphinium nuttallii, including within-population variation. Of the six cpDNA regions examined, five amplified reliably (22.1 kb total). Of the sixteen polymorphisms observed, all were restriction site mutations except for two length mutations. These resolved nine haplotypes, seven of which were unique to a single population (Figure 3). Three populations were polymorphic. Two haplotypes, B and D, were widespread. A single most-parsimonious network was found with one homoplastic trait (Figure 4). The AMOVA indicated a high level of genetic structuring (F ST = 0.87, P < 0.00001). In the nested contingency analysis, the null hypothesis of no geographical association was rejected for all but the highest clade level (P < 0.05). In the nested clade analysis, the nesting diagram contained three two-step clades (Figure 5). Patterns at the total cladogram level were explained by a contiguous range expansion, associated with the clade of western haplotypes (Figure 6). At the two-step clade level, contiguous range expansion or long-distance colonization were indicated. Past fragmentation or isolation by distance were cited at all one-step clade levels. Ambiguity at one- and two-step clade levels could not be resolved because of insufficient sampling of intermediate populations.
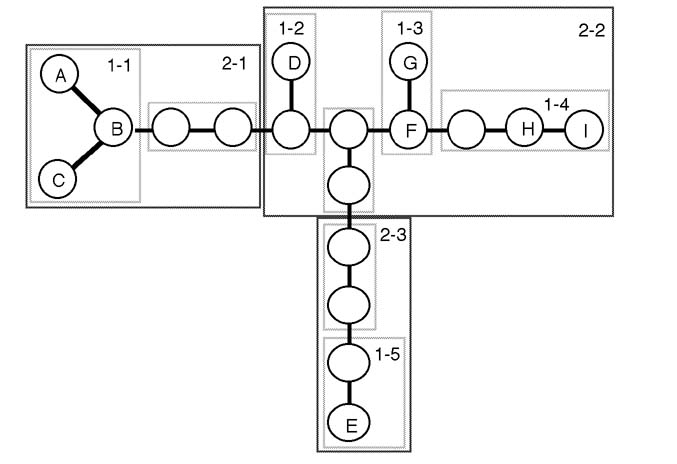
Figure 5. Nesting diagram for chloroplast DNA haplotype network. |

Figure 6. Nested clade analysis results and inferences. Terms explained in Templeton et al (1995). |
DISCUSSION & CONCLUSIONS
Significant genetic structuring was detected in D. nuttallii as well as within-population variation for chloroplast DNA markers. Significant geographical associations were detected by the exact nested contingency analysis, but results did not specify the source of these associations. AMOVA results suggested that, under equilibrium, observed patterns of genetic variation in D. nuttallii could be explained solely by limited dispersal. The low dispersal capability of the species supports this claim. In addition to isolation by distance, nested clade analysis uncovered several historical explanations for these patterns. While specific conclusions were limited by the scale of geographic sampling, there was little support for long-distance colonization or past fragmentation. At higher clade levels, a contiguous range expansion was identified associated with the western and northern portions of the species' current range. Movement from the eastern populations may have followed both the post-Pleistocene glacial retreat and the Missoula Floods in the Columbia River Gorge (15,000 - 13,500 years before present), with populations advancing as new habitat became available. Increased geographic sampling is underway for D. nuttallii to improve the resolution of the nested clade analysis. We predict a similar phylogeographic pattern might be observed for other plant species associated with Garry oak (Quercus garryanna), demonstrating a post-Pleistocene contiguous range expansion.REFERENCES
1. Templeton AR, Routman E, Phillips CA (1995) Separating population structure from population history: A cladistic analysis of the geographic distribution of mitochondrial DNA haplotypes in the tiger salamander, Ambystoma tigrinum. Genetics 140: 767-782.2. Templeton AR (1998) Nested clade analyses of phylogeographic data: testing hypotheses about gene flow and population history. Molecular Ecology 7: 381-397.
3. Beattie KL (1999) Interspecific and intraspecific genetic diversity in the chloroplast genome of Delphinium leucophaeum and D. nuttallii. BA thesis, Reed College.
4. Demesure B, Sodzi N, Petit RJ (1995) A set of universal primers for amplification of polymorphic non-coding regions of mitochondrial and chloroplast DNA in plants. Molecular Ecology 4: 129-131.
5. Dumolin-Lapegue S, Pemonge MH, Petit RJ (1997) An enlarged set of consensus primers for the study of organelle DNA in plants. Molecular Ecology 6: 393-397.
6. Templeton AR, Boerwinkle E, Sing CF (1987) A cladistic analysis of phenotypic associations with haplotypes inferred from restriction endonuclease mapping. I. Basic theory and an analysis of alcohol dehydrogenase activity in Drosophila. Genetics 117: 343-351.
7. Templeton AR, Sing CF (1993) A cladistic analysis of phenotypic associations with haplotypes inferred from restriction endonuclease mapping. IV. Nested analyses with cladogram uncertainty and recombination. Genetics 134: 659-669.
8. Posada D, Crandall KA, Templeton A (1999) GeoDis 2.0. Department of Zoology, Brigham Young University.
9. Schneider S, Roessli D, Excoffier L (2000) Arlequin ver. 2.000: A software for population genetics data analysis. Genetics and Biometry Laboratory, University of Geneva.